
timidin kinaz - Thymidine kinase
| timidin kinaz | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 U. urealyticum'dan bir timidin kinaz tetramerinin kristal yapısı ( monomerler sırasıyla renkli camgöbeği, yeşil, kırmızı ve macentadır) timidin ile kompleks halinde ( boşluk doldurma modeli , karbon = beyaz, oksijen = kırmızı, nitrojen = mavi) .
| |||||||||
| tanımlayıcılar | |||||||||
| AB numarası | 2.7.1.21 | ||||||||
| CAS Numarası. | 9002-06-6 | ||||||||
| veritabanları | |||||||||
| IntEnz | IntEnz görünümü | ||||||||
| BRENDA | BRENDA girişi | ||||||||
| ExPASy | NiceZyme görünümü | ||||||||
| fıçı | Fıçı girişi | ||||||||
| MetaCyc | metabolik yol | ||||||||
| PRİAM | profil | ||||||||
| PDB yapıları | RCSB PDB PDBe PDB toplamı | ||||||||
| Gen ontolojisi | AmiGO / QuickGO | ||||||||
| |||||||||
| timidin kinaz | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| tanımlayıcılar | |||||||||||
| Sembol | TK | ||||||||||
| Pfam | PF00265 | ||||||||||
| Pfam klanı | CL0023 | ||||||||||
| InterPro | IPR001267 | ||||||||||
| PROZİT | PDOC00524 | ||||||||||
| |||||||||||
| Timidin kinaz 1, çözünür | |||||||
|---|---|---|---|---|---|---|---|
| tanımlayıcılar | |||||||
| Sembol | TK1 | ||||||
| NCBI geni | 7083 | ||||||
| HGNC | 11830 | ||||||
| OMIM | 188300 | ||||||
| Referans Sırası | NM_003258 | ||||||
| UniProt | P04183 | ||||||
| Diğer veri | |||||||
| EC numarası | 2.7.1.21 | ||||||
| yer | Chr. 17 q23.2-25.3 | ||||||
| |||||||
| Timidin kinaz 2, mitokondriyal | |||||||
|---|---|---|---|---|---|---|---|
| tanımlayıcılar | |||||||
| Sembol | TK2 | ||||||
| NCBI geni | 7084 | ||||||
| HGNC | 11831 | ||||||
| OMIM | 188250 | ||||||
| Referans Sırası | NM_004614 | ||||||
| UniProt | O00142 | ||||||
| Diğer veri | |||||||
| EC numarası | 2.7.1.21 | ||||||
| yer | Chr. 16 [1] | ||||||
| |||||||
Timidin kinaz bir enzimdir , bir fosfotransferaz (bir kinaz): 2'-deoksitimidin kinaz, ATP-timidin 5'-fosfotransferaz, EC 2.7.1.21. Çoğu canlı hücrede bulunabilir. Memeli hücrelerinde TK1 ve TK2 olmak üzere iki formda bulunur. Bazı virüsler ayrıca viral timidin kinazların ekspresyonu için genetik bilgiye sahiptir. Timidin kinaz reaksiyonu katalize eder:
- Thd + ATP → TMP + ADP
Thd'nin (deoksi) timidin olduğu , ATP'nin adenosin trifosfat olduğu , TMP'nin (deoksi) timidin monofosfat olduğu ve ADP'nin adenosin difosfat olduğu . Timidin kinazlar, DNA sentezinde ve dolayısıyla hücre bölünmesinde anahtar bir işleve sahiptir , çünkü bunlar, timidin'i DNA'ya sokan benzersiz reaksiyon zincirinin bir parçasıdır. Timidin, gıdalardan ve ölü hücrelerden DNA'nın bozulmasının bir sonucu olarak vücut sıvılarında bulunur. Birçok antiviral ilacın etkisi için timidin kinaz gereklidir . Monoklonal antikorların üretiminde hibridoma hücre dizilerini seçmek için kullanılır . Olarak , klinik kimya , bir şekilde kullanılan çoğalma marker tanı , tedavi ve takibinde kontrolü malign hastalık esas olarak, hematolojik maligniteler .
Tarih
Timidinin DNA'ya dahil edilmesi 1950 civarında gösterildi. Bir süre sonra, bu birleştirmenin fosforilasyondan önce geldiği gösterildi ve 1960 civarında sorumlu enzim saflaştırıldı ve karakterize edildi.
sınıflandırma
İki farklı timidin kinaz sınıfı tanımlanmıştır ve bu süper aileye dahil edilmiştir: bir aile, herpes virüsünden timidin kinazı ve ayrıca hücresel timidilat kinazları birlikte gruplandırır, ikinci aile grupları, omurgalılar , bakteriler , bakteriyofaj T4'ü içeren çeşitli kaynaklardan TK. , poksvirüsler , Afrika domuz ateşi virüsü (ASFV) ve Balık Lymphocystis hastalığı virüsü (FLDV). Böcek yanardöner virüslerin ana kapsid proteini de bu aileye aittir. Prosite modeli, timidin kinazların yalnızca hücresel tipini tanır.
izozimler
Memelilerin kimyasal olarak çok farklı iki izoenzimleri vardır: TK1 ve TK2. İlki ilk önce fetal dokuda bulundu, ikincisinin yetişkin dokusunda daha bol olduğu bulundu ve başlangıçta bunlar fetal ve yetişkin timidin kinaz olarak adlandırıldı. Yakında TK1'in sitoplazmada sadece hücre bölünmesi beklentisiyle (hücre döngüsüne bağlı) mevcut olduğu, TK2'nin mitokondride bulunduğu ve hücre döngüsünden bağımsız olduğu gösterildi. İki izoenzim farklı reaksiyon kinetiğine sahiptir ve farklı inhibitörler tarafından inhibe edilir.
Viral timidin kinazlar hem yapısal hem de biyokimyasal olarak memeli enzimlerinden tamamen farklıdır ve memeli enzimlerini inhibe etmeyen inhibitörler tarafından inhibe edilir. İki insan izoenziminin genleri 1970'lerin ortalarında lokalize edildi. TK1 geni klonlandı ve dizilendi. Karşılık gelen proteinin moleküler ağırlığı yaklaşık 25 kD'dir. Normalde dokuda moleküler ağırlığı yaklaşık 50 kD olan bir dimer olarak bulunur. ATP tarafından aktive edilebilir. Aktivasyondan sonra, moleküler ağırlığı 100 kD civarında olan bir tetramerdir. Bununla birlikte, dolaşımda bulunan enzimin formu, gen tarafından kodlanan proteine karşılık gelmez: Dolaşımdaki aktif enzimin ana fraksiyonu 730 kD'lik bir moleküler ağırlığa sahiptir ve muhtemelen diğer proteinlere bir kompleks içinde bağlanır. Bu kompleks daha kararlıdır ve daha düşük moleküler ağırlıklı formların herhangi birinden daha yüksek bir spesifik aktiviteye sahiptir.
Rekombinant TK1, bu şekilde aktive edilemez ve bir tetramere dönüştürülemez, bu da hücrelerde meydana gelen enzimin sentezden sonra modifiye edildiğini gösterir.
TK1, hücre bölünmesinin S fazı sırasında hücre tarafından sentezlenir. Hücre bölünmesi tamamlandıktan sonra TK1 hücre içinde parçalanır ve normal hücre bölünmesinden sonra vücut sıvılarına geçmez. Hücrede timidin kinazın etkisinin bir geri besleme düzenlemesi vardır: timidin kinazın daha fazla fosforilasyonunun ürünü olan timidin trifosfat (TTP), timidin kinaz için bir inhibitör görevi görür . Bu, sistemi aşırı doyurmadan nükleik asit sentezi için mevcut dengeli bir TTP miktarının korunmasına hizmet eder. Timidinin toksik olmayan bir analoğu olan 5'-aminotimidin, bu düzenleyici mekanizmaya müdahale eder ve böylece antineoplastik ilaçlar olarak kullanılan timidin analoglarının sitotoksisitesini arttırır . Reaksiyon kinetiği timidin ve timidin analogları fosforilasyon karmaşık ve kısmen bilinmektedir. Timidinin timidin trifosfata genel fosforilasyonu Michaelis-Menten kinetiğini takip etmez ve timidin ve üridinin çeşitli fosfatları birbirlerinin fosforilasyonuna müdahale eder. Farklı türlerden gelen TK kinetikleri birbirinden farklıdır ve ayrıca belirli bir türün farklı formları (monomer, dimer, tetramer ve serum formu) farklı kinetik özelliklere sahiptir.
Virüse özgü timidin kinazlar için genler, Herpes simpleks virüsü, Varicella zoster virüsü ve Epstein-Barr virüsünde tanımlanmıştır.
 + ATP --->
+ ATP --->
 + ADP
+ ADP
Timidin, timidin monofosfat ve ADP vermek üzere ATP ile reaksiyona girer.
İşlev
Timidin kinaz tarafından katalize edilen reaksiyonun ürünü olan timidin monofosfat, sırasıyla timidilat kinaz enzimi tarafından timidin difosfata ve ayrıca enzim nükleozid difosfat kinaz tarafından timidin trifosfata fosforile edilir . Trifosfat, bir DNA polimeraz ve tamamlayıcı bir DNA molekülü (veya ters transkriptaz durumunda bir RNA molekülü , retrovirüste bulunan bir enzim ) tarafından katalize edilen bir reaksiyon olan bir DNA molekülüne dahil edilir .
Timidin monofosfat da farklı bir reaksiyon içerisinde hücrenin ürettiği metilasyon bir deoksiüridin monofosfat enzimi, timidin ile ilgisi olmayan diğer metabolik yolların bir ürünü, timidilat sintaz . İkinci yol, DNA onarımı için timidin monofosfat sağlamak için yeterlidir. Bir hücre bölünmeye hazırlandığında, eksiksiz bir yeni DNA kurulumu gerekir ve timidin trifosfat da dahil olmak üzere yapı taşlarına olan gereksinim artar. Hücreler, bölünme sırasında gerekli olan bazı enzimleri yaparak hücre bölünmesine hazırlanırlar. Normalde hücrelerde bulunmazlar ve daha sonra aşağı regüle edilirler ve bozulurlar . Bu tür enzimlere kurtarma enzimleri denir. Timidin kinaz 1 böyle bir kurtarma enzimidir, oysa timidin kinaz 2 ve timidilat sentaz hücre döngüsüne bağlı değildir.
Eksiklik
Timidin kinaz 2, hücreler tarafından mitokondriyal DNA sentezi için kullanılır. TK2 genindeki mutasyonlar , mitokondriyal DNA tükenme sendromunun miyopatik bir formuna yol açar . TK 2 eksikliğinin bir başka nedeni de oksidatif stres kaynaklı S-glutationilasyon ve mitokondriyal timidin kinaz 2'nin proteolitik yıkımı olabilir . TK1 eksikliğinin neden olduğu hiçbir sendrom bilinmemektedir, muhtemelen kusurlu bir TK1 geni fetüsün ölümüne yol açacaktır.
Gelişim sırasında timidin kinaz
Sentezden sonra timidin kinaz 1'in modifikasyonundan sonra tetramer oluşumu, enzim aktivitesini arttırır. Bunun enzim aktivitesinin düzenlenmesi için bir mekanizma olduğu öne sürülmüştür. Dictyostelium gelişim aşamasından sonra tetramerlerin oluşumu gözlenir. DNA sentezinin ince düzenlenmesi için kullanımının, sıcak kanlı hayvanlarda omurgalılardan ayrıldıktan sonra kurulduğu ileri sürülmektedir. Ayrıca gelişimde timidin kinaz benzeri enzimlerin gelişimi de incelenmiştir.
Tür dağılımı
Timidin kinaz hayvanlarda, bitkilerde, bazı bakterilerde, arkelerde ve virüslerde bulunur. Pox virüslerinden, Afrika domuz ateşi virüsünden, Herpes simpleks virüsünden, Varicella zoster virüsünden ve Epstein-Barr virüsünden timidin kinazlar tanımlanmış ve değişen derecelerde karakterize edilmiştir. Virüsteki enzim formu diğer organizmalardakinden farklıdır. Mantarlarda timidin kinaz bulunmaz .
Uygulamalar
Bölünen hücrelerin tanımlanması
Biyokimyasal araştırmalarda timidin kinazın ilk dolaylı kullanımı, bölünen hücrelerin radyoetiketli timidin eklenmesiyle tanımlanması ve ardından bölünen hücreleri tanımlamak için radyoaktivite veya otoradyografinin ölçülmesiydi. Bu amaçla tritiye edilmiş timidin, büyüme ortamına dahil edilir. Teknikteki hatalara rağmen, malign hücrelerin büyüme oranını belirlemek ve immünolojide lenfositlerin aktivasyonunu incelemek için hala kullanılmaktadır.
Aktif tümörlerin PET taraması
Florotimidin bir timidin analoğudur . Alımı timidin kinaz 1 tarafından düzenlenir ve bu nedenle tercihen hızla çoğalan tümör dokusu tarafından alınır. Flor izotopu 18, pozitron emisyon tomografisinde (PET) kullanılan bir pozitron yayıcıdır . Flor-18 radyoetiketli florotimidin F-18 bu nedenle aktif tümör proliferasyonunun PET görüntülemesi için yararlıdır ve daha yaygın olarak kullanılan işaretçi florodeoksiglukoz (18F) ile olumlu şekilde karşılaştırılır . Klinik çalışmaların karşılaştırılmasına yardımcı olacak standartlaştırılmış bir protokol önerilmiştir.
Hibridomların seçimi
Hibridomalar , (sonsuz sayıda bölünebilen) tümör hücreleri ile immünoglobulin üreten lenfositlerin (plazma hücreleri) kaynaştırılmasıyla elde edilen hücrelerdir. Hibridomalar, belirli bir benzersiz spesifikliğe (monoklonal antikorlar) sahip büyük miktarlarda immünoglobulinler üretecek şekilde genişletilebilir. Bir problem, hücre füzyonundan sonra büyük miktarda kaynaşmamış hücreden hibridomları ayırmaktır. Bunu çözmenin yaygın bir yolu , füzyon için timidin kinaz negatif (TK-) tümör hücre dizilerini kullanmaktır. Timidin kinaz negatif hücreler, timidin kinaz pozitif (TK+) hücreleri öldüren timidin analoglarının varlığında tümör hücre çizgisinin büyütülmesiyle elde edilir. Negatif hücreler daha sonra genişletilebilir ve TK+ plazma hücreleri ile füzyon için kullanılabilir. Füzyondan sonra hücreler, dihidrofolat redüktaz enzimini inhibe eden ve böylece timidin monofosfatın de novo sentezini bloke eden metotreksat veya aminopterin içeren bir ortamda büyütülür. Yaygın olarak kullanılan bu tür bir ortam, hipoksantin, aminopterin ve timidin içeren HAT ortamıdır. Timidin kinaz eksikliği olan hücre dizisinden gelen kaynaşmamış hücreler, timidin monofosfat kaynağına sahip olmadıkları için ölürler. Lenfositler, "ölümsüz" olmadıkları için sonunda ölürler. Sadece hücre hattı atalarından "ölümsüzlüğe" sahip hibridomlar ve plazma hücresinden timidin kinaz hayatta kalır. Arzu edilen antikoru üretenler daha sonra seçilir ve monoklonal antikoru üretmek için kültürlenir. Hibridoma hücreleri, kurtarma yolunda GMP nükleotit sentezi için gerekli IMP'yi sentezleyen HGPRT adlı başka bir gen ile ilgili olarak tarif edilenle aynı prensip kullanılarak da izole edilebilir.
Kromozom yapısının incelenmesi
DNA liflerinin moleküler taranması, tomurcuklanan Saccharomyces cerevisiae mayasındaki kromozomların yapısını izlemek için kullanılabilir. Bu, bireysel moleküllerin DNA replikasyon profillerini sağlar. Bu, maya suşlarının timidin kinazı eksprese etmesini gerektirir, ki bu yabani tip mayalarda bulunmaz, mantarlar (bkz. oluşum). Bu nedenle, timidin kinaz için bir gen, genoma dahil edilmelidir.
klinik kimya
Timidin kinaz, yalnızca hücre bölünmesi beklentisiyle mevcut olan bir kurtarma enzimidir. Enzim, hücrelerin, hücre bölünmesinden sonra artık ihtiyaç duymayan proteinleri parçalamak için özel bir mekanizmaya sahip olduğu normal bölünme geçiren hücrelerden serbest bırakılmaz. Normal kişilerde, serum veya plazmadaki timidin kinaz miktarı bu nedenle çok düşüktür. Tümör hücreleri, muhtemelen ölü veya ölmekte olan tümör hücrelerinin bozulmasıyla bağlantılı olarak dolaşıma enzim salmaktadır. Bu nedenle serumdaki timidin kinaz seviyesi, dolaylı olarak tümörün saldırganlığının bir ölçüsü olarak, kötü huylu çoğalmanın bir ölçüsü olarak hizmet eder.
Terapötik uygulamalar
Bazı ilaçlar özellikle bölünen hücrelere yöneliktir. Hastalıklı hücreler normal hücrelerden çok daha sık çoğaldıkları için tümörlere ve viral hastalıklara (hem retrovirüse hem de diğer virüslere karşı) ve ayrıca aşırı hızlı hücre replikasyonu ile ilgili bazı habis olmayan hastalıklara (örneğin sedef hastalığı) karşı kullanılabilirler. Timidin analoglarının antiviral ve anti-kanser aktivitesinin, en azından kısmen, mitokondriyal timidin kinazın aşağı regülasyonu ile elde edildiği öne sürülmüştür.
sitostatik
Timidin metabolizmasına karşı yönlendirilen ve dolayısıyla kanserle ilişkili hücre bölünmesini kontrol etmek için kullanılan timidin kinazı içeren farklı ilaç sınıfları vardır. Zincir sonlandırıcılar, büyüyen DNA zincirine dahil olan ancak zincirin daha fazla uzatılamayacağı şekilde modifiye edilen timidin analoglarıdır. Timidinin analogları olarak, bu tip ilaçlar kolaylıkla 5'-monofosfatlara fosforile edilir. Monofosfat ayrıca ilgili trifosfata fosforile edilir ve büyüyen DNA zincirine dahil edilir. Analog, devam eden zincir büyümesi için gerekli olan 3'-pozisyonunda hidroksil grubuna sahip olmayacak şekilde modifiye edilmiştir. Zidovudin'de (AZT; ATC:J05AF01) 3'-hidroksil grubu bir azido grubu ile değiştirildi, stavudin'de (ATC: J05AF04) değiştirilmeden çıkarıldı. AZT, serumda timidin kinaz belirleme yöntemlerinden birinde substrat olarak kullanılır. Bu, AZT'nin bu yöntemi engellediği ve bir sınırlama olabileceği anlamına gelir: AZT, HIV enfeksiyonunda HAART tedavisinin standart bir bileşenidir. AIDS'in yaygın bir sonucu lenfomadır ve timidin kinaz tayininin en önemli tanısal uygulaması lenfomanın izlenmesi içindir.

AZT
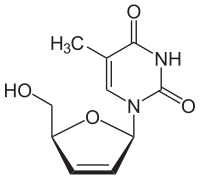
stavudin

idoksuridin

asiklovir
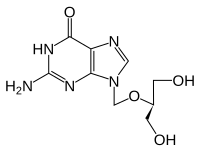
gansiklovir





Diğer timidin analogları , örneğin İdoksuridin (ATC: J05AB02), müteakip replikasyon döngüleri sırasında baz eşleşmesini bloke ederek etki eder, böylece ortaya çıkan DNA zincirini kusurlu hale getirir. Bu aynı zamanda, malign hücrelerin apoptozunu sağlamak için radyoaktivite ile birleştirilebilir.
antiviraller
Asiklovir (ATC: J05AB01) ve gansiklovir (ATC: J05AB06) gibi bazı antiviral ilaçlar ve diğer nükleozid analogları, insan timidin kinazlarının aksine viral timidin kinazın substrat özgüllüğünü kullanır. Bu ilaçlar, kendi içlerinde toksik olmayan ancak viral timidin kinaz tarafından fosforilasyon yoluyla toksik ilaçlara dönüştürülen ön ilaçlar olarak işlev görür. Bu nedenle virüsle enfekte olan hücreler, hücre ölümüne yol açan oldukça toksik trifosfatlar üretirler. İnsan timidin kinazı, aksine, daha dar özgüllüğü ile ön ilacı fosforile edemez ve aktive edemez. Bu şekilde, yalnızca virüs tarafından enfekte olan hücreler ilaca duyarlı hale gelir. Bu tür ilaçlar, yalnızca spesifik timidin kinazları ile herpes grubundan virüslere karşı etkilidir. Bu tip ilaçlarla tedavi edilen hastalarda antiviral ilaç direnci gelişimi sıklıkla gözlenir. Herpes simpleks virüsü ve Varicella zoster virüsünde timidin kinaz geninin dizilenmesi, hızlı genetik değişkenliği gösterir ve antiviral ilaç direnci tanısını kolaylaştırabilir.
Aralık 1979'da DSÖ tarafından çiçek hastalığının eradike edildiğinin ilan edilmesinden sonra aşılama programları sonlandırıldı. Hastalığın tesadüfen veya biyolojik savaş sonucu yeniden ortaya çıkması, korunmasız bir popülasyonla karşılaşacak ve kontrolü zor olabilecek bir salgına neden olabilecektir. Çiçek hastalığı salgınıyla mücadele etmek için toplu aşılama zor olabilir çünkü onaylanmış tek çiçek hastalığı aşısı olan Vaccinia Virus'un ciddi yan etkileri olabilir. Bununla birlikte, bazı hükümetler, olasılığa karşı sigortalamak için Çiçek aşısı stokluyorlar. Bununla birlikte, spesifik ve etkili antiviral ilaçların geliştirilmesine öncelik verilmektedir. Olası bir yaklaşım, poksvirüsün timidin kinazının özgüllüğünü, herpes virüsüne karşı ilaçlar için kullanıldığı şekilde, amaç için kullanmak olabilir. Bir zorluk, poksvirüs timidin kinazının, insan timidin kinazları ile aynı timidin kinaz ailesine ait olması ve dolayısıyla kimyasal olarak daha benzer olmasıdır. Poksvirüs timidin kinazlarının yapısı bu nedenle potansiyel antiviral ilaçları bulmak için belirlenmiştir. Ancak araştırma henüz çiçek virüslerine karşı kullanılabilir bir antiviral ilaçla sonuçlanmadı.
Gen tedavisinde “intihar geni” olarak
Herpes virüsü timidin kinaz geni, gen terapisi deneylerinde bir güvenlik sistemi olarak bir "intihar geni" olarak da kullanılmış ve geni ifade eden hücrelerin gansiklovir kullanılarak öldürülmesine olanak sağlamıştır. Bu, rekombinant genin kontrolsüz hücre büyümesine (insersiyonel mutagenez) yol açan bir mutasyona neden olması durumunda arzu edilir. Bu modifiye edilmiş hücreler tarafından üretilen sitotoksik ürünler komşu hücrelere difüze olabilir ve bu onları benzer şekilde "seyirci etkisi" olarak bilinen bir fenomen olan gansiklovire duyarlı hale getirebilir. Bu yaklaşım, hayvan modellerinde kanseri tedavi etmek için kullanılmıştır ve tümörün, geni eksprese eden habis hücrelerin en az %10'u ile öldürülebilmesi açısından avantajlıdır. Benzer bir sistem, domates timidin kinaz ve AZT kullanılarak denenmiştir. Ek olarak, timidin kinaz geni, 2016 yılında Avrupa'da şartlı olarak onaylanan Zalmoxis adlı hematopoietik kök hücre nakli tedavisinde tehlikeli graft-versus-host hastalığıyla mücadele etmek için bir intihar geni olarak kullanılmaktadır.
Tümör belirteç genleri
Timidin kinazın benzer bir kullanımı, bazı tümör hücrelerinde normal hücrelerde bulunmayan maddelerin ( tümör belirteçleri ) varlığından yararlanır . Bu tür tümör belirteçleri, örneğin, CEA (karsinoembriyonik antijen) ve AFP'dir (alfa fetoprotein). Bu tümör markörleri için genler, timidin kinaz için promotör genler olarak kullanılabilir. Timidin kinaz daha sonra tümör markörünü eksprese eden hücrelerde aktive edilebilir, ancak normal hücrelerde değil, öyle ki gansiklovir ile tedavi sadece tümör hücrelerini öldürür. Bu tür gen terapisine dayalı yaklaşımlar, tümör hücrelerine gen transferinin hedeflenmesiyle ilgili problemler henüz tamamen çözülmediği için hala deneyseldir.
Tümörler için nötron yakalama tedavisi
Bir timidin analoğunun bor ile birleştirilmesi önerilmiş ve beyin tümörlerinin bor nötron yakalama tedavisi için hayvan modellerinde denenmiştir. Bor içeren çok sayıda timidin türevi tanımlanmıştır.
parazit önleyiciler
Bir parazit genomuna bir TK geninin dahil edilmesi, BrdU'nun dahil edilmesini mümkün kılar ve böylece paraziti bu ilaçla tedaviye duyarlı hale getirir ve ayrıca önerilmiştir ve parazit genomunun replikasyonunun hassas bir göstergesini oluşturur.
Ölçüm
Serum ve plazmada
Serum veya plazmadaki timidin kinaz seviyeleri çoğunlukla enzim aktivite deneyleri kullanılarak ölçülmüştür. Ticari deneylerde bu, bir serum numunesinin bir substrat analoğu ile inkübasyonu ve oluşan ürün miktarının ölçülmesiyle yapılır. Timidin kinaz proteininin immünoanaliz ile doğrudan belirlenmesi de kullanılmıştır. Bu yöntemle bulunan timidin kinaz miktarları, enzim aktiviteleriyle iyi bir korelasyon göstermez. Bunun bir nedeni, immünoassay ile tanımlanan büyük miktarda serum TK1'in enzimatik olarak aktif olmamasıdır. Bu, özellikle immünoanalizlerin daha duyarlı olabileceği katı tümörler için geçerlidir.
dokuda
Timidin kinaz, dokunun ekstraksiyonundan sonra doku örneklerinde belirlendi. Ekstraksiyon veya tahlil için standart bir yöntem geliştirilmemiştir ve hücrelerden ve dokulardan alınan ekstraktlarda TK tayini herhangi bir spesifik klinik soruyla ilgili olarak doğrulanmamıştır, ancak bkz. Romain ve ark. ve Arner et al. Substrat analogu 5-Bromovinil 2'-deoksiüridin kullanılarak hücre özütlerinde TK2'nin spesifik tespiti için bir yöntem geliştirilmiştir. Aşağıda belirtilen çalışmalarda kullanılan yöntemler ve sonuçların raporlanma şekli o kadar farklıdır ki, farklı çalışmalar arasında karşılaştırma yapmak mümkün değildir. Gelişim sırasında fetal dokulardaki TK1 seviyeleri, daha sonra karşılık gelen dokularınkinden daha yüksektir. Bazı habis olmayan hastalıklar da hücrelerde ve dokularda TK değerlerinin çarpıcı bir şekilde yükselmesine neden olur: monositoz sırasında periferik lenfositlerde ve pernisiyöz anemi sırasında kemik iliğinde. TK1 hücre bölünmesi sırasında hücrelerde mevcut olduğundan, malign dokudaki TK aktivitesinin karşılık gelen normal dokudan daha yüksek olması gerektiğini varsaymak mantıklıdır. Bu, çoğu çalışmada da doğrulanmaktadır.
immünohistokimyasal boyama
İmmünohistokimyasal tespit için timidin kinaza karşı antikorlar mevcuttur. Timidin kinaz için boyama, evre 2 meme karsinomlu hastaların tanımlanmasında güvenilir bir teknik olarak bulundu. Tespit edilen en yüksek hasta sayısı, timidin kinaz ve Ki-67 boyamasının kombinasyonu ile elde edilmiştir. Teknik ayrıca akciğer kanseri, kolorektal karsinom, akciğer kanseri ve renal hücreli karsinom için doğrulanmıştır.
Floresan boyama
2'-deoksi-2',2'-difloro-5-etiniluridin (dF-EdU) Herpes simpleks virüsü timidin kinaza bağlanır, ancak sterik engel nedeniyle insan timidin kinazına bağlanmaz. Bu reaktif, bir floresan azit ile birlikte, enfekte olmuş hücrelerin floresansına neden olur, ancak enfekte olmayan hücrelerde değil. Bu nedenle, bu substrat analoğu, enfekte olmuş hücreleri spesifik olarak boyamayı mümkün kılar.
Ayrıca bakınız
- Timidin kinaz 1
- Herpes virüsünden timidin kinaz
- timidilat kinaz
- nükleosit-difosfat kinaz
- timidilat sentaz
- Klinik kimyada timidin kinaz
Referanslar
daha fazla okuma
- Garone C, Garcia-Diaz B, Emmanuele V, Lopez LC, Tadesse S, Akman HO, et al. (Ağustos 2014). "Timidin kinaz 2 eksikliği için deoksipirimidin monofosfat baypas tedavisi" . EMBO Moleküler Tıp . 6 (8): 1016–27. doi : 10.15252/emmm.201404092 . PMC 4154130 . PMID 24968719 .
- O'Neill KL, Buckwalter MR, Murray BK (Kasım 2001). "Timidin kinaz: tanısal ve prognostik potansiyel". Moleküler Teşhisin Uzman İncelemesi . 1 (4): 428–33. doi : 10.1586/14737159.1.4.428 . PMID 11901857 . S2CID 12790327 .
- Topolcan O, Holubec L (Şubat 2008). "Kanser hastalıklarında timidin kinazın rolü". Tıbbi Teşhis Konusunda Uzman Görüşü . 2 (2): 129–41. doi : 10.1517/17530059.2.2.129 . PMID 23485133 .
- Jagarlamudi KK, Shaw M (Eylül 2018). "Bir tümör biyobelirteç olarak timidin kinaz 1: teknik gelişmeler eski bir biyobelirteç için yeni bir potansiyel sunuyor". Tıpta Biyobelirteçler . 12 (9): 1035–1048. doi : 10.2217/bmm-2018-0157 . PMID 30039979 .
Dış bağlantılar
- ABD Ulusal Tıp Kütüphanesi Tıbbi Konu Başlıklarında (MeSH) Timidin+kinaz